

by Cody Haun, PhD (c), MA, CSCS
& Brandon Roberts, PhD, CSCS
Before reading this, you may want to take a look at Part 1 for a refresher on concurrent training. As we delve into Part 2, we will cover the molecular exercise physiology, then provide some application for concurrent programming. Our goal is to describe some of the more important underlying mechanisms involved in the acute responses and chronic adaptive processes. We intend to explain how interference can occur, to what extent, and why this matters for programming applications. At the end, we provide an example of how concurrent training can be applied within the context of a mesocycle of training. Let’s get started with:
Factors that contribute to the interference effect acutely and chronically:
- Molecular signaling
- Muscle fiber type
- Muscle glycogen
- Time between bouts of endurance and resistance training
Factor #1: Molecular Signaling
Hypertrophy is the product of muscle protein breakdown (MPB) and muscle protein synthesis (MPS). Two of the most well-established stimuli of muscle hypertrophy are: 1) resistance training, and 2) protein consumption. Interestingly, Part 1 introduced the idea that concurrent training may produce greater muscle hypertrophy than resistance training alone in some contexts. This runs counter to industry dogma and other evidence suggesting an interference effect. If the interference effect is true, then at least some physiological evidence or rationale must exist to underpin this concept. In 2014, Dr. Keith Baar published an excellent paper titled, “Using Molecular Biology to Maximize Concurrent Training.”
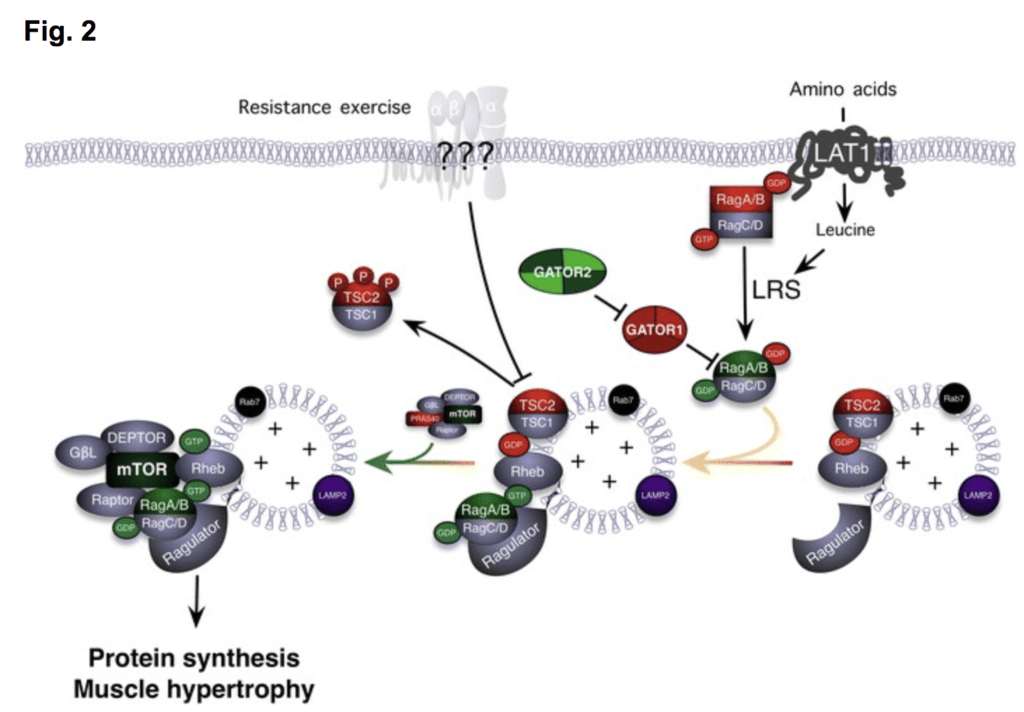
Resistance exercise stimulates signaling of various proteins at the membrane (protein structures shown near membrane with “???” transposed) of muscle via the sensing of tension, which results in downstream signaling. One of these signaling proteins, mTOR, can result in significant elevations of MPS. Certain other signaling proteins are stimulated from endurance exercise (e.g., AMPK, SIRT1), and these molecules can transiently inhibit mTOR complex formation, which can ultimately affect the magnitude of MPS after training.
For hypertrophy to occur, the synthesis of muscle proteins must outpace the breakdown of muscle protein. Naturally, certain signaling processes precede the synthesis of a muscle protein. mTOR is a protein in skeletal muscle that is currently recognized as a central convergence point of a variety of anabolic signals. As proof of concept, studies have shown that when mTOR signaling is inhibited via delivery of a drug (i.e., rapamycin) after resistance exercise, MPS is blunted.
We are particularly interested in how endurance training may interfere with the mTOR signaling pathway. Interference is currently thought to primarily occur through an increase in the activity of another signaling protein called adenosine monophosphate kinase (AMPK). AMPK activity increases in muscle cells when AMP begins to accumulate, due to the increased breakdown of ATP for energy during muscle contraction. Since absolute ATP turnover is higher during repetitive contraction (like endurance exercise), evidence suggests that AMPK may be a key interference signaling molecule generated from endurance exercise. An image from John Hawley elegantly demonstrates this idea.
If stimulated too close to resistance training, AMPK is thought to be one explanatory mechanism of the interference effect. However, as noted by Dr. Baar, this explanation doesn’t completely explain interference in humans. Rather, other signaling factors related to metabolic stress caused by the accumulation of free radicals, lactate, calcium, and — particularly — a family of signaling proteins known as NAD+-dependent deacetylases (e.g., SIRT1) can also influence interference.
Two studies from Dr. John Hawley’s laboratory demonstrate a dose-dependent interference effect beautifully. In the first of the two studies, 10 six-second, all-out cycling sprints were performed 15 minutes prior to or after completing 8 sets of 5 repetitions at 80% 1RM. Completing these all-out cycling sprints prior to resistance training completely blunted the typical increase in mTOR signaling after resistance training, whereas, mTOR signaling was comparatively higher if these sprints were completed after resistance training. In a follow-up study where the intensity of the conditioning was significantly reduced prior to or after resistance training (~70% VO2 max) and the duration only lasted 30 minutes, Dr. Hawley’s group showed that the interference effect on mTOR signaling didn’t occur, regardless of if conditioning was performed before or after resistance training.
Therefore, low- to moderate-intensity conditioning does not seem to exert this effect, but, if performed close to resistance training, is likely best placed after a resistance training bout. A recent systematic review concluded that placing weights before conditioning, if performed in proximity to one another, resulted in, on average, a 4kg higher squat 1RM after a training intervention when compared to subjects performing endurance training before lifting. The authors concluded: “Sequencing strength training prior to endurance in concurrent training appears to be beneficial for lower body strength adaptations….”.
To summarize, one of the most well-established molecular signaling explanations of the interference effect is related to the impaired upregulation of the Pi3K-Akt-mTORc1 pathway. Since endurance exercise activates AMPK and SIRT1, which may inhibit mTOR, this may suppress muscle protein synthesis (MPS). AMPK and SIRT1 activity levels seem to return to baseline activity levels relatively more quickly after endurance exercise versus more prolonged increases in mTOR signaling activity after resistance exercise (3h versus 18h). However, at least four studies suggest that concurrent training neither hinders or benefits myofibrillar MPS. Although beyond the scope of this article, specific genetic, nutritional, and other individual factors are probable to explain a fair share of the variation in individual responses.
Factor #2: Muscle Fiber Types
Conventional fiber-typing of muscle tissue (i.e., fast versus slow) indicates acute molecular signaling differences from training. In general, motor neurons (and the muscle fibers they innervate) are recruited based on their size. Primarily fast-twitch muscle fibers are recruited during bouts of higher-intensity training or training involving near-maximal or maximal efforts, given their contractile properties and the size of the motor neurons innervating these fibers. Consequently, firing of the motor neurons that innervate these fast-twitch fibers is unlikely to result from bouts of lower-intensity, steady-state conditioning to a meaningful extent. So, the molecular signaling phenomena related to MPS discussed above is therefore unlikely to occur in these larger, predominantly fast-twitch fibers from bouts of lower-intensity, steady-state conditioning since primarily slow-twitch fibers are sufficient to produce the forces necessary for continued running or low-intensity, repetitive work.
Evidence suggests a short-term increase in muscle size from concurrent training likely due to the increase in contractile activity of slower-twitch fibers, at least in the thigh. This seems logical, since the knee extensors are involved in bouts of extended duration running and since the quadriceps are a “mixed” group of fiber-types containing ~50% predominantly slow-twitch fibers. The observed increase in muscle size of the quadriceps in studies involving concurrent training is potentially due to the relative increase in recruitment of slow-twitch fibers of the thigh with hypertrophy occurring as a consequence.
Resistance training alone at a high intensity recruits both slow-twitch fibers and fast-twitch fibers, but slow-twitch fibers are recruited at an absolutely lower extent when calculated relative to time, compared to adding in a few hours of running per week in addition to resistance training. To put it in perspective: Resistance training at a high intensity recruits both fast- and slow-twitch muscle fibers (e.g., the quads during a heavy squat); however, the absolute amount of time slow-twitch fibers experience tension is lower compared to if you were running a few times a week. Since muscle fiber contraction exists on an all-or-none continuum, the relative increase in the recruitment of slow-twitch fibers to contract and generate tension across their length in a cycle of concurrent training seems to increase the growth response of these fibers. In comparison, recruitment of these fibers, on an absolute scale at least, during periods of resistance training alone hypothetically provides a lower growth stimulus to the fibers on a mesocycle scale.
Factor #3: Glycogen
Glycogen is an important substrate in skeletal muscle that allows the synthesis of ATP which can be used to provide the energy for muscle contraction. The use of glycogen is primarily determined by the intensity and duration of exercise. As exercise intensity increases, skeletal muscle begins to shift to a greater proportion of carbohydrate oxidation, with muscle glycogen being the primary source. In fact, the liver only stores ~100g of glycogen compared to the ~300-700g in skeletal muscle in most humans (glycogen amount is variable based on diet, training level, sex, body weight, and other factors).
In muscle, glycogen is distributed within the fiber in three ways: subsarcolemmal (~10-15%), intermyofibrillar (~70-80%), and intramyofibrillar (~10-15%).
The location of glycogen is key as each of these glycogen depots seem to play different roles. Intermyofibrillar glycogen helps power calcium signaling important to the release of calcium for muscle contraction, and appropriate reuptake for release thereof for continued contractile ability. Intramyofibrillar glycogen seems to more directly power cross-bridge cycling and is preferentially depleted during HIIT. In particular reference to why glycogen is being discussed in this article, glycogen depletion is highly correlated with fatigue during exercise (Knuiman et al., 2017).
In one study using a glycogen depletion protocol, strength was decreased in an isoinertial task, but not isokinetic. Isoinertial is the more important of the two in regards to translation to powerlifting training since it mimics normal training, whereas isokinetic training is completed at a constant speed and is thus ecologically invalid for the powerlifter, in general. Therefore, although relatively low glycogen levels may not significantly hamper productive training in a single powerlifting training session, some evidence indicates that it may. Additionally, enough evidence exists suggesting a performance benefit from more full glycogen stores persuades us to recommend making an effort to defend muscle glycogen stores in context of intense powerlifting training. To provide a bit of peace of mind if your carbohydrate intake wasn’t on point in a given day in which you are lifting, in 2012, Camera et al. showed that resistance training in a relatively low glycogen state didn’t seem to reduce muscle protein synthesis significantly, at least when measured acutely (Camera et al., 2012). Hence, although glycogen levels are certainly of relevant concern to the individual seeking to maximize training performance and the adaptive response thereof, the extent to which these levels impact powerlifting training and adaptation likely matters somewhat less in comparison to trainees focused more on maximizing muscle hypertrophy.
When we deplete glycogen, exercise performance can be impaired even though other fuel sources are available (e.g., fat, protein). However, a typical strength training session only reduces glycogen levels by ~20-40%, depending mainly on the amount of volume completed. Unlike bodybuilders, who tend to complete more total volume, powerlifters generally train with lower volumes and higher intensities, so they may not reduce glycogen as much during a training session; although, this amount can vary depending on the phase of training the lifter is in. Practically, some of the potentially negative effects of concurrent training on resistance training adaptations due to alterations in glycogen stores are likely of greater concern for individuals training with higher volumes.
Factor #4: Time between bouts of endurance and resistance training
There are two primary hypotheses to explain an interference effect between endurance and resistance exercise in reference to time: 1) the acute hypothesis, and 2) the chronic hypothesis.
The acute hypothesis suggests that it is the short-term change in the magnitude of muscle damage or glycogen depletion from either type of training that transiently affects the other type of training. This is thought to primarily occur through the hampered ability to train during the successive session at a requisite intensity, or with a requisite volume, to facilitate an improvement in muscle size or strength. Practically speaking, this hypothesis, if true, indicates that small but meaningful reductions in training intensity or volume due to residual fatigue from proximal bouts of training on the same day or within the same week accumulate over enough time to equal out to a meaningful decrement in progress.
The chronic hypothesis contends that skeletal muscle can’t adapt metabolically and morphologically to both resistance training and endurance training within the same program of sufficient duration (i.e., ~6-10 weeks), given that select adaptive patterns to either type of training are rather distinct (e.g., changes in enzyme content).
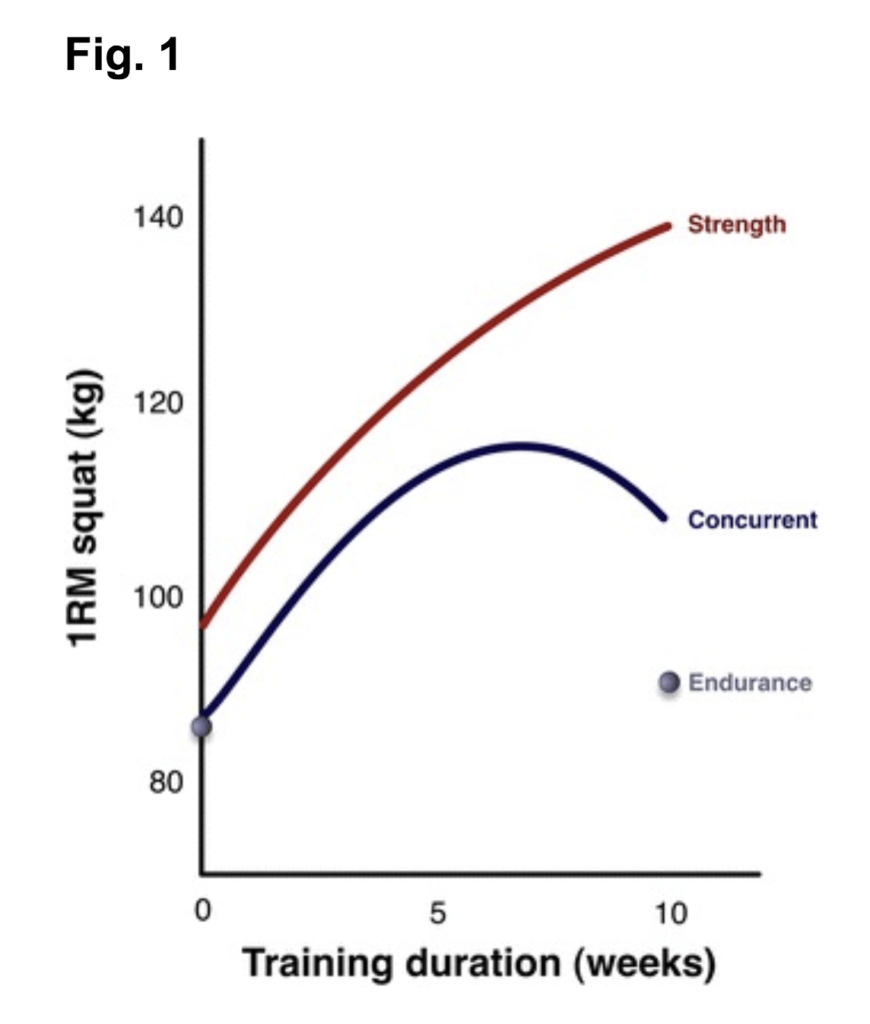
We think it is most likely a combination of both acute and chronic factors. Chronic alterations in a phenotype or performance are necessarily related to the summation of the acute responses (i.e., specificity). With this in mind, a fascinating figure from Dr. Lawrence Spriet’s laboratory helps clarify the pattern of adaptive effects shown in the figure below.
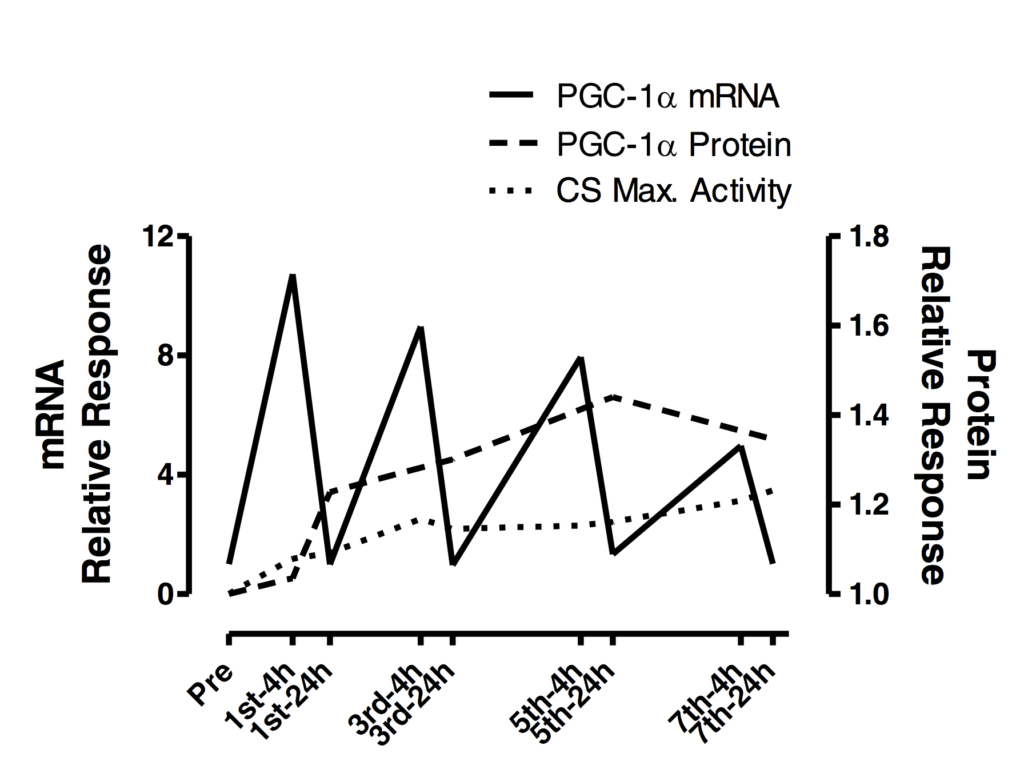
In the figure above, the amount of a protein related to aerobic metabolism gradually increases to a point after two weeks of almost 50% more protein in the muscle than when the training began. In light of this trend, we think that the observed interfering changes over time between the two modes begins to clearly present only when enough time spent training with either mode results in an accumulation of various metabolic and structural proteins resulting in a phenotype. At 24h, the mRNA shown above is back to baseline, while the protein amount is still gradually increasing. Although an oversimplification, placing a bout of resistance training at this point is likely to stimulate a distinct transcriptional and translational response, and inhibit the translational events shown above to a lesser extent compared to placing this resistance training bout in closer proximity to the measured mRNA peak.
In order to examine the effects of endurance training, we have to analyze the data from the time of training to the time of the next training session to determine if there is a detrimental effect via impairments in work capacity, strength, or both. In an extreme example of this: One study had subjects complete a 5km run, then tested maximal repetitions performed in four sets of 80% 1RM in the half squat after 30 minutes, 60 minutes, 4 hours, 8 hours, and 24 hours. The important finding from this study was that a significant drop in total volume only occurred at 30 and 60 minutes post-endurance training. Using a less extreme example, another study compared concurrent training with high-intensity interval training (HIIT) (6 x 3 mins @ 95-100% effort) or submaximal training (36 minutes at 70% effort). There was no difference between the modality of endurance training, but the total volume of strength training was found to be lower at 4h and 8h, while it recovered by 24 hours.
More recently, completion of HIIT in hopes of mitigating interference effects has received attention. An additional benefit posited by HIIT proponents is that HIIT takes much less time to complete than submaximal endurance training. A study used HIIT training 0h, 6h, or 24h after strength training and found the 6h or 24h response to be similar. Those who completed HIIT directly after strength training (0h) didn’t improve nearly as much in strength outcomes such as bench press and half squat over a period of seven weeks (Robineau et al., 2017).
On the other hand, a study found that HIIT (10 sets of 60 seconds with 100% effort with 60s between bouts) directly after heavy resistance exercise did not inhibit significant strength or hypertrophy after 8 weeks of training, but rather promoted cardiovascular and endurance adaptations (Tsitkanou et al., 2016). Upon closer look, the group who did not concurrently train had a +9% improvement in strength from baseline. If we look at a few more percentages, the concurrent group had a larger increase in size of the vastus lateralis, yet the strength group had a larger increase in the rectus femoris (i.e., regional hypertrophy). There was also a seldom-mentioned detriment in the group that performed concurrent training – a lower rate of force development, which might be important for powerlifters (Tsitkanou et al., 2016).
Application
Factor #1 Application (Molecular Signaling)
If muscle hypertrophy or strength is the primary pursuit of training, conditioning performed around resistance training at a very high intensity can interfere with the adaptive response to resistance training. Training modalities are likely best separated by at least three hours. Preferably, we advise separating them by ~24 hours for insurance, but we will describe why in more detail below.
Factor #2 Application (Fiber type, Bioenergetic, and Systemic Considerations)
While we can’t exclusively target specific muscle fiber types in a practical sense, or control our inherited percentages, there are more systemic benefits of low-intensity conditioning practices worthy of mention. Namely, an increased ability of the cardiovascular system to transport and deliver oxygen-rich blood to muscle tissue between sets and sessions of resistance training can benefit the powerlifter. The benefits of a modest degree of cardiovascular fitness for the serious powerlifter can translate to quicker recovery between sets or training sessions, and therefore the opportunity to present an overload through training that could be missed with sub-par levels of cardiovascular fitness. Since this point seems vague, we’d like to qualify it a bit more in relation to the interaction between systemic and local factors.
Many of you are likely familiar with the term “bioenergetics.” This term refers to the process by which organisms use energy to produce work. Regarding muscle physiology, muscle cells (i.e., fibers) utilize three categorical divisions of bioenergetics, based on their relative contribution to work over time: a) immediate (i.e., ATP-PCr), b) glycolytic (discussed above and below), and c) aerobic or oxidative mechanisms. To be brief, these categories are concomitantly active, but to a lesser or greater extent depending on the intensity and duration of exercise. They are ordered above in terms of their relative contribution to producing ATP to perform muscular work over time. To be clear, in terms of time: 1) the ATP-PCr “system” is primarily active during the first 10-20 seconds of muscular work, 2) the glycolytic system is primarily active between 20-120 seconds (depending on the number of glycogen stores, or the size of glycogen granules in a muscle fiber), and 3) the oxidative system, or mitochondrial respiration, is primarily active thereafter. These time frames are simply rounded values and are not absolute, depending on a variety of factors. Regardless, this information is commonly employed to refute the importance of aerobic metabolism (i.e., mitochondrial activity) in relation to powerlifting training, since most powerlifting training occurs in the range of <= ~20 seconds. However, in our opinion, it is vital to the experienced trainee, or coach, to understand how PCr is resynthesized to appreciate the importance of the oxidative system in muscle cells as this pertains to powerlifting programming. To be concise, PCr is primarily resynthesized through the provision of ATP via aerobic metabolism. Perhaps the visual below is helpful to drive this point home.
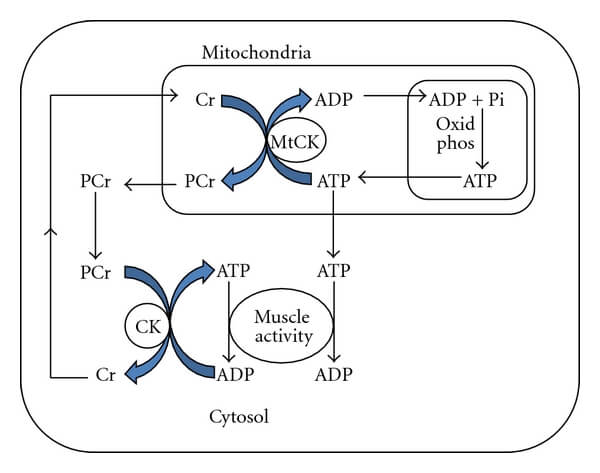
This image demonstrates the relationship between oxidative phosphorylation (ADP + Pi → ATP) and PCr resynthesis (ATP + Cr → PCr). ATP, produced from oxidative phosphorylation, can interact with an enzyme, creatine kinase, which catalyzes the linking of a phosphate group to creatine (Cr above) resulting in PCr. Why does this matter? Well, how quickly and to what extent PCr is resynthesized between sets is at least cursorily related to how effectively oxygen is delivered to muscle fibers between sets and the ability of muscle tissue to transport and utilize this oxygen for the provision of ATP to resynthesize PCr for high-intensity muscle contraction. Clearly, this is important for the production of force during heavy sets of 1-5 reps, since the immediate energy system is the primary system active. That is, the systemic factors related to the transportation of oxygen-enriched blood and local ability of the muscle fibers reduced of PCr during intense muscular work are intricately related and can be rate-limiting in the sense of supplying the ATP important for producing maximum forces in successive sets of powerlifting training. OK, OK, enough of the mumbo-jumbo, how does this pertain to programming? All other things being equal, the powerlifter with greater cardiovascular abilities to transport oxygen, consume oxygen locally (at the muscle fiber level), and utilize this oxygen to resynthesize ATP for PCr resynthesis between sets can theoretically recover faster between sets to produce higher forces within a whole session involving multiple sets. Consequently, this individual can present a more powerful overloading stimulus through training, albeit with a number of assumptions being made. In a sense, this can be considered a greater acute recoverability.
This translates to a greater propensity to provide a more powerful stimulus during overloading sessions of training for the lifter with greater cardiovascular fitness, assuming no decrement in muscle mass, neural, or structural interference through too much concurrent, or endurance training. This means adaptations from a modest amount of endurance training or low-intensity, more aerobic metabolism-based work can translate to quicker recovery between sets. Although the means to this end can look quite different, practically, adaptations like: a) an increased stroke volume (blood pumped per heartbeat), b) greater perfusion of muscle fibers reduced of PCr during intense contraction (increased capillarization of muscle tissue), and c) an increased mitochondrial density in muscle fibers or increased oxidative enzyme content can improve recovery between sets. These are hallmark adaptations of endurance training. Furthermore, glycolytic activity provides upstream signals to increase mitochondrial activity, which are important for this process to occur. Hence, even in context of more voluminous resistance training (e.g., sets of >6 reps), this argument still holds water since glycolytic activity increases as a function of PCr breakdown, and this feeds into increased oxidative metabolism. To summarize, a practical method of increasing the ability to perform muscular work during a resistance training session aiming to increase maximum strength is to improve the ability of the lifter’s cardiovascular system, and local muscle ability, to transport and utilize oxygen for the resynthesis of ATP. Enter, appropriate dosing of conditioning not directly interfering with resistance training adaptations (i.e., sound concurrent training), at least at some points within the macrocycle of training.
Factor #3 Application [Glycogen]
Each type of training (i.e., endurance and resistance) should alternate foci between the primary musculature used during each session, in reference to completed training the day before or the day after. This is to avoid any potential interference in training performance if glycogen stores are reduced to suboptimal levels in muscle used in the successive session. For example, this could mean placing upper body-based conditioning bouts in closer proximity to lower body resistance training or placing lower body-based conditioning bouts in closer proximity to upper body resistance training.
Factor #4 Application [Time between bouts]
Positioning each type of training on, ideally, separate days altogether (~24h) or at least a few hours after one another could help avoid interference. Since residual fatigue from endurance training can potentially reduce the tension developed during the strength portion of training, care should be taken to allow enough time for the perception of fatigue from either bout to dissipate.
Programming Examples
With a tip of the hat to periodized training, we show an example of two microcycles. In this case, seven days of training for a powerlifter about 6 months away from a meet. Since peaking is of no real concern in the short-term, these microcycles are snapshots of: microcycle 1 of a 4-microcycle length mesocycle, which includes 3 overloading micros, followed by 1 deloading microcycle. Also, microcycle 3 is shown as an example of a functional overreaching microcycle which precedes the deloading microcycle. The overarching goal of this mesocycle is to increase work capacity.
Since all fitness characteristics (e.g., strength, power, speed, etc.) have unique decay rates, a certain dose of conditioning will be required to maintain proximal levels of cardiovascular fitness in upcoming training cycles. However, the dose of training to maintain a characteristic is not the same as the dose to improve a characteristic. Consequently, the session parameters and programming structure will, and should, be different in upcoming cycles wherein the focus will be based on the specific peaking date of the lifter. Some of the basic session parameters of individual sessions are provided (note that conditioning sessions are ~24 hours away from all weight training sessions).
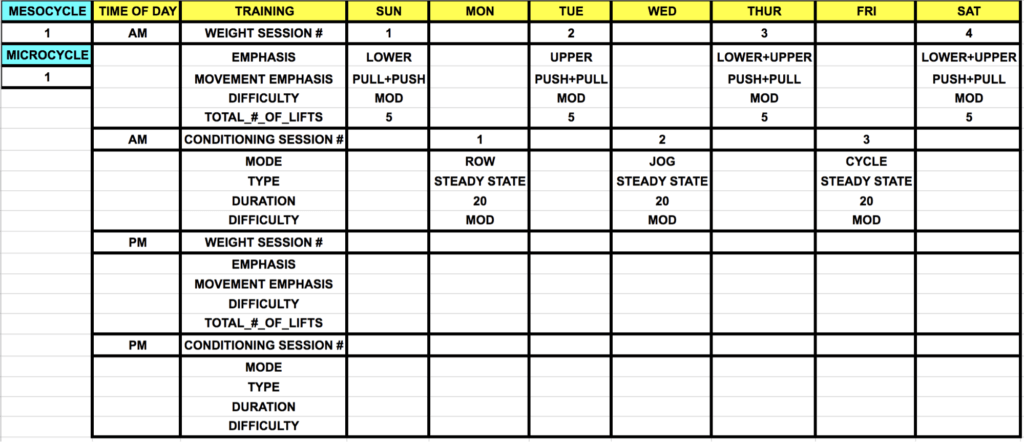
“MOD” in the above example stands for moderate difficulty. For weight sessions, on average, this refers to a relative intensity of the session sufficient to stimulate an adaptive process, but not so aggressive to overreach too early in the cycle. Therefore, most lifts will be performed a few repetitions shy of technical failure. Weight sessions will be performed at ~5 reps per set, and at an intensity corresponding to ~75% absolute 1RM of a movement. “PULL” and “PUSH” indicate the type of movement to be primarily performed in the session, and the order of listing is intentional (i.e., PULL+PUSH versus PUSH+PULL). PULL+PUSH indicates that both lower body pulling and pushing movements will be performed, but pulling exercises (e.g., deadlift, row) will precede pushing exercises (e.g., Squat, OH Press). Similarly, the order of LOWER+UPPER indicates the primary emphasis of the session. As you can see, the conditioning mode and movement pattern of the mode is positioned in such a way to avoid interference with the successive training bout.
Regarding conditioning sessions, a moderate difficulty refers to an intensity corresponding to ~70-75% maximum heart rate (220-age), and therefore a % of VO2 max around 70%. Hence, neither the weight training nor the conditioning is inherently too difficult the first microcycle in way of providing an overall greater adaptive stimulus over the course of the mesocycle. We advise caution in the earlier microcycles to avoid the accumulation of fatigue impairing the ability to productively train in later microcycles. Keep in mind, programming this type of training is as much an art as it is a science.
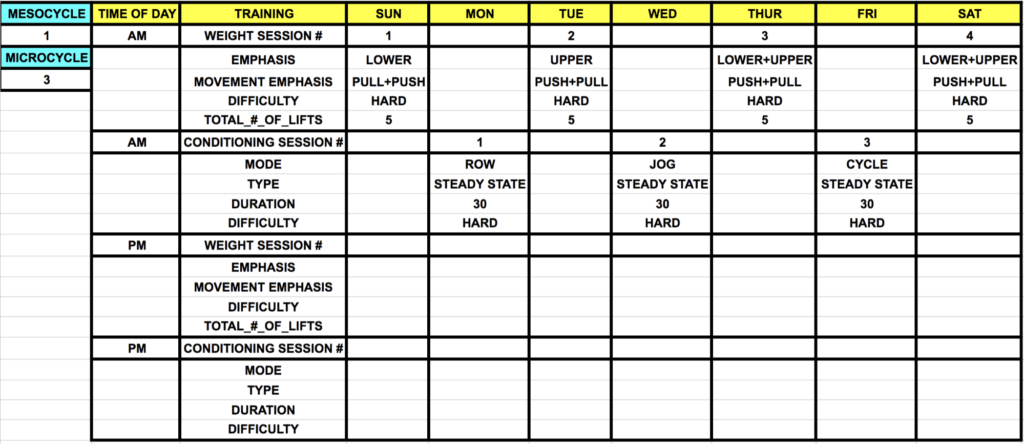
In this case, the absolute intensity of each working set in the weight room will have increased to ~85-90% 1RM, and sets will be more proximal to failure (i.e., ~1 rep shy of technical failure). Conditioning session difficulty will approach ~75-85% VO2max, and a % of maximum HR corresponding to ~85%. In the case of concurrent training in context of seeking to build muscle or increase strength, a simplified strategy to avoiding combative signaling is to separate training modes by at least 24 hours. This provides assurance, based on the molecular signaling and applied work discussed above that the adaptive “area under the curve” is almost entirely realized from any given bout before an additional stimulus is applied.
In summary:
- If necessary to complete in a crunch for time, conditioning should be completed after resistance training and not before.
- If concurrent training is employed in a powerlifting context, positioning more voluminous blocks relatively far away from peaking blocks is a good idea.
- During blocks of training focused on improving work capacity wherein concurrent training may be undertaken, separating endurance and resistance training bouts by ~24 hours provides insurance for avoiding any interference with heavy weight training.
- Placing conditioning modalities that stimulate different musculature than immediately adjacent resistance training sessions (i.e., the day before or the day after) provides further assurance that interference won’t occur.
- If conditioning modes are completed within a few hours of resistance training, these bouts should be more moderate or low in intensity.
Feel free to shoot us an email for any specific questions you may have regarding this information or if you are interested in receiving personal training programming services designed for your specific goals.
Cody Haun, PhD(c), MA, CSCS
Email: codythaun@gmail.com
Webpage: www.strongerbyscience.com/coach-cody
Brandon Roberts, PhD, MS, CSCS
Email: robertsb21@gmail.com
Webpage: www.thestrengthguys.com
Read Next
- Concurrent Training for the Powerlifting, Part 1: Introduction and Background
- Avoiding Cardio Could be Holding You Back


